제아잔틴에 의한 RAW264.7 세포에서의 항염효과
요약
목적
본 연구는 제아잔틴에 의한 RAW264.7 세포의 항염 효과를 확인하기 위하여 수행되었다.
방법
제아잔틴이 RAW264.7세포에 항염효과를 나타내는지 확인하기 위하여, 제아잔틴 에 의한 세포생존률 측정, 싸이토카인의 유전자 발현양상, PGE2와 산화질소의 생성 정도를 확인하였다.
결과
본 연구를 통해 제아잔틴이 iNOS, COX2, 인터류킨6, TNFα 유전자의 발현을 감소시킴을 확인할 수 있었다. 또한 산화질소의 생성을 감소시키고 PGE2의 생성을 감소시킴을 통해 제아잔틴의 항염효과를 확인할 수 있었다.
결론
본 연구를 통해 제아잔틴의 항염효과를 확인하였고, 제아잔틴이 항염 및 아토피를 타겟으로 하는 화장품 원료로서의 가능성을 제시하였다. 제아잔틴이 피부미용, 화장품, 건강식품 등의 산업 등에도 응용될 수 있을 것으로 여겨지며 그 가능을 확인하기 위해 제아잔틴이 염증세포에 미치는 기전연구가 필요할 것으로 보인다.
핵심용어: 제아잔틴, 항염, 산화질소, PGE2, 인터류킨
Abstract
Purpose
This study was performed to test the anti-inflammatory effects of zeaxanthin on skin using RAW264.7 cells.
Methods
The anti-inflammatory effects of zeaxanthin on RAW264.7 cells were assessed by measuring cell viability, mRNA expression, and nitric oxide (NO)/prostaglandin E2 (PGE2).
Results
The anti-inflammatory effects of zeaxanthin were elucidated by analysis of IL1α/IL1β/IL6/TNFα mRNA expression and NO/PGE2 production. Quantitative real-time polymerase chain reaction showed that zeaxanthin decreased the mRNA level of IL6 and TNFα. Additionally, PGE2/NO detection also revealed that zeaxanthin exhibited anti-inflammatory properties.
Conclusion
The results presented in this study suggest that zeaxanthin is an anti-inflammatory compound. Therefore, it could be a potent cosmetic ingredient with anti-inflammatory effects against atopic dermatitis. Further research on the anti-inflammatory mechanisms of zeaxanthin would not only facilitate the development of functional cosmetics but also pharmacological treatments.
Keywords: Zeaxanthin, Anti-inflammation, PGE2, Nitric oxide, Interleukin
中文摘要
目的
探索玉米黄质对RAW264.7细胞的皮肤的抗炎作用。
方法
通过测量细胞活力,mRNA表达和一氧化氮(NO)/前列腺素E2(PGE2)来评估玉米黄质对RAW264.7细胞的抗炎作用。
结果
通过分析IL1α/IL1β/IL6/TNFαmRNA表达和NO/PGE2产生来阐明玉米黄质的抗炎作用。实时定量聚合酶链反应显示玉米黄质降低了IL6和TNFα的mRNA水平。此外,PGE2/NO检测还显示了玉米黄质具有抗炎特性。
结论
这项研究提出的结果表明玉米黄质是一种抗炎化合物。因此,它可能是一种有效的化妆品成分,对特应性皮炎具有抗炎作用。玉米黄质的抗炎机制的进一步研究不仅将促进功能化妆品的开发,而且还将促进药物治疗。
关键词: 玉米黄质, 抗炎, 一氧化氮, PGE2, 白介素
Introduction
염증은 외부 자극원, 손상된 세포 및 박테리아 등을 포함한 유해한 생체 내외부 자극에 대한 면역 반응으로 알려져 있다( Albina & Reichner, 1995). 즉, 생명체의 세포 및 조직을 보호하기 위한 면역반응이지만 염증이 과도하게 유발되면 당뇨병, 고혈압, 아테로마성 동맥경화증, 암 및 신경 퇴행성 장애 등과 같은 질환 등을 통해 세포 및 조직 손상을 유도 할 수 있다고 알려져 있다( Albina & Reichner, 1995). 세균의 세포벽을 구성하고 있는 lipopolysaccharide (LPS)는 대식세포(macrophage)의 Toll-like receptor (TLR)에 결합하여 세포 내부 신호전달경로인 nuclear factor kappa beta (NFĸB)와 mitogen-activated protein kinase (MAPK) 신호의 활성화를 유도하고, prostaglandin E2 (PGE 2), nitric oxide (NO)와 같은 염증성 매개인자를 분비하고 interleukin-1 (IL1), interleukin-6 (IL6), tumor necrosis factor-α (TNFα)와 같은 염증성 싸이토카인 분비를 증가시킨다( Akira & Takeda, 2004; Gomez et al., 2005). 대식세포는 면역 반응에서 여러 생물학적 과정에 중요한 역할을 하는 세포로써, 면역조절, 항원 제시, 식균작용과 같은 세 가지 주요 기능을 담당하며 이를 위해 다양한 싸이토카인 및 염증매개인자를 방출한다( Akira & Takeda, 2004; Gomez et al., 2005). 이렇게 생성된 염증 매개인자들은 주변세포를 보호하거나 면역기능을 강화시키는 중요한 역할을 하기도 하지만 과도한 NO와 PGE 2 등은 염증을 악화시키는 원인이 되기도 한다( Kim et al., 2004). 특별히 NO는 염증의 대표적 지표물질로, 일반적으로 NO는 종양을 억제하거나 박테리아 등의 미생물을 죽이는 중요한 역할을 하지만 과도하게 생성된 NO는 유전적 변이, 조직손상 및 신경손상 등을 유발한다( Gabay, 2006; van Triel et al., 2010; Lee et al., 2014; Im, 2014). 또한 cyclooxygenase 2 (COX2)는 arachidonic acid을 염증매개물질인 prostaglandin E2 (PGE 2)로 생성하는 효소로서 iNOS 등과 함께 염증반응에 중요한 유전자로 알려져 있다. LPS에 의해 대식세포가 활성화되면 NO와 PGE 2는 의 생성이 증가되고 이는 IL1, 6 및 TNFα와 같은 염증성 싸이토카인 등을 생산한다( Horwood et al., 2006; Hirohashi & Morrison, 1996). 따라서 NO나 PGE 2, 염증성 싸이토카인 등의 생성억제는 염증성 질환을 예방이나 치료에 있어 중요한 목표가 된다. 카로티노이드(carotenoids)는 널리 분포된 화합물의 일종으로 노란색, 오렌지색, 붉은색 등을 띄는 천연 색소의 일종이다. 이러한 색소들은 세균, 조류, 균류, 식물 등에 의해 합성되고 식품첨가물, 건강보조식품, 가금류 및 동물의 조사료 등으로 사용되고 있다. 카로티노이드는 강력한 항산화( Bartley & Scolnik, 1995; Kurihara et al., 2002) 및 항암효과( Mayne, 1996; Bertram et al., 2005), 면역기능 향상( Amar et al., 2004; Chew & Park, 2004) 등의 다양한 생물학적 효능을 지닌다고 알려져 있다. 특별히 파프리카, 사프란, 구기자 등의 녹황색 채소에 풍부하게 존재한다고 알려진 제아잔틴(zeaxanthin [(3R,3R)-β,β-carotene-3,3diol])은 크산토필(xanthophyll)의 일종으로, 눈 건강에 매우 효과적인 소재로 알려져 있고 눈건강 보조제의 첨가제로 사용되고 있다( Berry et al., 2003; Harker et al., 1998; Hannibal et al., 2000). 하지만 제아잔틴이 RAW264.7 세포주에서의 염증 완화에 효과적인지에 대한 연구는 보고된 바가 없다. 본 연구에서는 LPS에 의해 자극이 발생된 RAW264.7 세포주에 제아잔틴을 처리하였을 때의 항염증 효과를 확인하였다. 제아잔틴에 의한 RAW264.7 세포주의 생존률, 대식세포 표지유전자의 발현과 NO, PGE2 단백질 발현을 CCK-8 실험법, Real-time RT-PCR 및 ELISA 실험법으로 각각 확인하였다.
Methods
1. RAW264.7 세포주 배양
RAW264.7 대식세포주는 한국세포주은행(Korean Cell Line Bank, Korea)에서 구매하여 사용하였다. 소태아혈청(Fetal bovine serum; FBS, Welgene, Korea)과 penicillin/streptomycin (Invitrogen, USA)이 첨가된 Dulbecco's modified Eagle's medium (DMEM; Gibco, USA)을 RAW264.7 대식세포주 배양에 사용했다. RAW264.7 대식세포주는 5 % CO2, 37℃, 100 % 습윤조건의 인큐베이터에서 배양하였다.
2. 세포 생존율 검증
제아잔틴(Cayman chemicals, USA)에 대한 세포의 생존율은 CCK-8 (Cell Counting Kit-8; Dojindo, Japan)를 사용하여 측정하였다. RAW264.7 대식세포주를 조직 배양 접시에 접종하고 24 h 동안 안정화 시킨 후, 제아잔틴을 농도 별로 24 h 처리하였다. 이후 phenol-red 무첨가 DMEM (Welgene)에 CCK-8 시약을 1:10의 비율로 희석하여 인큐베이터에서 30 min 동안 반응시켰다. Microplate reader (BioTek, USA)를 사용하여 450 nm 파장에서 흡광도를 측정하였고, 세포가 없는 배지를 대조군의 흡광도를 기초로 하여 세포 생존율을 계산하였다. 세포의 생존율은 대조군에 대한 백분율로 표시하였다.
3. 실시간 유전자 중합효소 연쇄반응(real-time RT-PCR)
Invitrogen의 TRIzol ® Reagent (USA)를 사용하여 RAW264.7 대식세포주의 total RNA를 추출하였다. 추출한 RNA로부터 SuperiorScriptTM III reverse-transcriptase (Enzynomics, Korea)를 이용하여 cDNA를 합성하였으며, RAW264.7 대식세포주 표지인자의 발현양상을 확인하기 위하여 StepOnePlus real-time PCR machine (Applied Biosystems, USA)을 이용하여 real-time RT-PCR을 진행하였다. 실험에 사용된 Taqman ® Gene expression assay (Applied Biosystems)는 Table 1에 명기하였다.
4. NO 정량
RAW264.7 대식세포주를 35 mm 조직 배양 접시에 20×104 cells로 접종하고 24 h 동안 안정화 시킨 후, 100 ng/mL 농도의 lipopolysaccharide (LPS; Sigma, USA)와 제아잔틴을 농도 별로 첨가하여 24 h 동안 배양 후, 상층액을 수집하여 원심분리한 후 상등액 100 μL를 이용하여 NO 생성량을 NO detection kit (iNtRON, Korea)를 사용하여 제조사의 프로토콜을 기반으로 측정하였다. 프로토콜을 간략히 요약하면, 100 μL 의 상등액에 50 μL의 N1 buffer를 첨가하여 10 min간 상온에서 반응시키고 N2 buffer를 50 μL 추가하여 10 min 간 추가로 반응시킨 후 ELISA reader로 520-560 nm의 파장에서 흡광도를 측정하였다.
5. PGE2-ELISA (Enzyme-linked immunosorbent assay)
제아잔틴이 PGE2의 발현에 끼치는 영향을 알아보기 위하여 RAW264.7 대식세포주를 24 h 동안 인큐베이터에서 안정화시킨 후, 100 ng/mL의 LPS와 제아잔틴을 농도 별로 첨가하여 6 h 동안 인큐베이터에서 배양하였다. PGE2-ELISA kit (Pierce Endogen, USA)를 사용하여 제조사의 프로토콜을 따라 RAW264.7 대식세포주가 분비하는 배양액 내의 PGE2 분비량을 측정하였다.
6. 통계분석
통계처리는 Student's t-test법을 이용하여 유의수준 0.05 이하(p<0.05)로 하여 검정하였다.
Results & Discussion
1. 제아잔틴의 농도 별 세포 생존율 분석
본 실험에 사용된 마우스 유래 대식세포주인 RAW264.7 대식세포주는, lipopolysaccharide (LPS)에 반응하여 염증반응이 나타나 염증성 싸이토카인을 방출하는 것으로 알려져 있다. 대식세포는 생체 전반에 분포하고 있으며 염증유발인자에 즉각적인 방어를 제공하는 면역세포로, 후천성 면역을 유도하는 항원제시세포로서의 역할과 다양한 싸이토카인을 분비하는 세포로 알려져 있다( Lawrence et al., 2002). 제아잔틴의 항염증 효과를 확인하기에 앞서, 연구에 사용하고자 하는 제아잔틴의 독성여부를 확인하기 위하여 CCK-8 assay로 세포의 생존률을 확인하였다. 대조군은 시료를 처리하지 않았고 제아잔틴은 100, 10, 1 μg/mL, 100, 10, 1, 0.1 ng/mL의 농도로 각각 처리하여 세포 생존율을 측정하였다( Figure 1). 100 μg/mL 농도 이상의 제아잔틴을 처리한 RAW264.7 대식세포주의 생존율이 유의성 있게 감소하였고, 10 μg/mL 이하의 농도에서는 RAW264.7 대식세포주의 생존율이 대조군과 유사하였기에 본 실험에서 진행되는 추가적인 실험은 10.1 μg/mL, 100 ng/mL의 제아잔틴을 처리하여 실험을 진행하였다.
2. 제아잔틴이 NO 생성에 미치는 영향
생체 내에서 NO는 혈압을 조절하거나, 종양과 세균을 제거하고 신호전달을 매개하는 등 다양한 역할을 담당하는 중요한 인자로 알려져 있다( Lee et al., 2014). 하지만 과도한 수준으로 생성되는 NO는 신경 등의 조직의 손상을 야기하고, 유전자의 변이를 유도하거나, 부종을 유발하는 등의 염증반응을 유도한다( Gabay, 2006; van Triel et al., 2010; Im, 2014). 본 실험에서 제아잔틴의 NO 감소 효과를 확인한 결과, LPS에 의해 11.5 μM 농도로 생성된 NO가 1 μg/mL의 제아잔틴을 처리하였을 때 5.8 μM로 나타나, LPS 처리군 대비 NO 생성이 유의성 있게 감소한 것으로 나타났다( Figure 2A). LPS는 대식세포에서 inducible NO synthase (iNOS)라는 NO생성을 유도하는 효소를 촉진하여 NO의 생성을 증가시킨다( Namazi, 2004). 따라서 NO를 생성하는 효소인 iNOS 유전자 발현을 제아잔틴이 감소시키는지 확인하기 위하여 quantitative Real-time RT-PCR을 수행하였다. iNOS의 발현이 대조군 대비 LPS 처리군에서 현저히 증가하였으나, 1 μg/mL의 제아잔틴을 처리한 RAW 264.7 세포주에서는 LPS 처리군 대비 유의성 있게 감소함을 확인하였다( Figure 2B).
3. 제아잔틴이 PGE2의 생성에 미치는 영향
Prostaglandin (PG)는 arachidonic acid에서 유래된 불포화지방산으로 염증반응을 유도하는 호르몬으로 알려져 있다( Yun et al., 2008). PG의 한 종류인 PGE 2는 염증반응을 매개하고, cyclooxygenase-2 (COX2) 효소에 의하여 생성된다. COX2 유전자가 과도하게 발현하거나 활성화된 경우에는 PGE 2의 생성이 증가되어 통증, 발열 및 염증반응이 야기된다( Harris et al., 2002; Namazi, 2004). 제아잔틴의 항염 효능을 확인하기 위하여 제아잔틴의 PGE 2 감소 여부를 확인한 결과, LPS 처리군에서는 PGE 2의 생성이 19.5 μg/mL로, 대조군에 비해 현저히 증가하였다. 그에 반해 1 μg/ml의 제아잔틴을 처리하였을 때에는 PGE 2의 생성이 6.3 μg/mL로 나타나 LPS 처리군과 비교하여 PGE 2 생성이 현저히 감소하였다( Figure 3A). 또한 PGE 2를 생성하는 효소인 COX2 유전자의 발현은 LPS 처리군에서 2.3배 증가하였으나 1 μg/mL의 제아잔틴을 처리한 세포주에서는 LPS 처리군 대비 42% 감소하였다( Figure 3B).
4. 제아잔틴이 염증성 싸이토카인의 생성에 미치는 영향
피부에의 만성적 염증은 피부의 노화 뿐만 아니라 피부질환을 야기할 수 있다( Noman et al., 2009; Shon et al., 2013; Lee et al., 2014). IL1α와 IL1β는 다양한 염증과정, 면역반응 및 조혈에 관여하는 싸이토카인으로, 대식세포와 면역반응이 유도된 형질세포 및 진피층내 섬유아세포 등에서 생성되는 염증의 강력한 매개체이다( Lord et al., 1991; Ban et al., 2011). 특히 IL1β는 염증부위에 호중구와 림프구 등의 염증세포의 침윤을 상승시키는 싸이토카인으로 잘 알려져 있다(Roitt et al., 2002; Baek et al., 2012). IL6는 형질세포의 분화와 성장을 유도하고, 알러지를 포함한 만성 염증성 질환을 유발한다고 보고되어 있다( Lee et al., 2014; Gabay, 2006). 또한, TNFα는 염증반응 초기에 염증부위로 호중구를 유도하여 급성 면역반응과 부종 및 발열을 유도하는 인자이다( Zhang et al., 1992). LPS는 RAW 264.7대식세포주에서 인터류킨류와 iNOS, TNFα 등의 염증매개 인자의 생산을 촉진한다. 본 연구에서는 LPS (100 ng/mL)를 처리하여 RAW 264.7대식세포주의 염증반응을 유도한 후 제아잔틴의 염증성 싸이토카인 감소여부를 확인한 결과, IL1α는 LPS 처리군에 의해 8.5배 증가하였으나 10 μg/mL의 제아잔틴을 처리하였을 때 오히려 LPS 처리군 대비 84% 가량 발현이 증가하였고 1 μg/mL과 100 ng/mL의 제아잔틴을 처리한 세포군에서는 LPS 처리군 대비 IL1α 유전자의 발현을 감소시키지 못했다( Figure 4A). IL1β는 LPS 처리군에 의해 3.8배 증가하였으나 제아잔틴에 의한 유전자 발현은 감소하지 않았다( Figure 4B). IL6는 1 μg/mL 의 제아잔틴을 처리하였을 때 LPS 처리군에 비하여 36% 가량 발현이 감소하였다( Figure 4C). TNFα 는 제아잔틴 1 μg/mL 의 농도에서 LPS 처리군에 비해 43% 가량 발현이 감소하였다( Figure 4D). 본 연구에서 제아잔틴이 염증에 관여하는 전사인자 중 IL6 와 TNFα 의 발현을 억제시켰으며, NO와 PGE 2 등의 생성 모두 1 μg/mL 농도의 제아잔틴 처리군에서 감소됨을 통하여 항염증 효과를 확인하였고, 피부노화 및 염증성 질환을 개선하는 치료제 또는 기능성 화장품 원료로서 제아잔틴을 활용할 수 있음을 보여주는 기초적인 자료가 될 것으로 보여진다. 특별히 Figure 2, 3와 Figure 4 C, D에서 1 μg/mL의 제아잔틴이 LPS에 의해 염증반응이 유도된 대식세포의 NO, PGE 2, IL6, TNFα의 생성 및 발현을 특이적으로 감소시키는 농도라고 보여지며, 화장품 등의 조성에 적절히 활용할 수 있을 것이다. 10 μg/mL 농도의 제아잔틴에 의해 iNOS/COX2 유전자의 발현이 감소하지 않고 그에 의한 NO/PGE 2 생성도 LPS 처리군과 비교하여 감소하지 않았다( Figure 2, 3). 그리고 IL1α, β 유전자의 발현은 오히려 10 μg/mL 농도의 제아잔틴 처리군에서 증가됨을 확인하였다( Figure 4A, B). 이는 CCK-8 assay를 통해서 10 μg/mL 농도의 제아잔틴이 세포생존율에 영향을 끼치지는 않지만 RAW264.7 세포주에 상당한 스트레스를 유발하는 농도로 여겨지며 소재로 사용될 시 적절한 농도를 배합하여 제품에 적용해야 할 필요성을 암시하고 있다.
Conclusion
본 연구에서 제아잔틴을 RAW264.7 대식세포에 처리할 적절한 농도를 확인할 수 있었다. 10 μg/mL 이하의 농도에서는 RAW264.7 대식세포주의 생존율에 영향을 미치지 않음을 Figure 1을 통하여 확인하였다. iNOS, COX2, IL6와 TNFα의 유전자 발현양상 측정과 PGE 2 단백질 발현 측정, NO 생성량 측정에서 제아잔틴이 유의성 있게 LPS에 의해 증가된 염증매개 인자의 발현이 회복되는 결과를 나타내었다( Figure 2- 4). 이는 제아잔틴이 새로운 피부 염증개선 후보물질로서의 가능성을 보여준다. 제아잔틴의 피부의 염증을 개선할 수 있는 가능성을 확인한 연구는 본 연구가 최초로, 추후 피부미용 혹은 피부질환 치료제로서 실질적으로 활용될 수 있고, 염증에 의한 피부노화를 예방할 수 있는 가능성을 제시한 결과라 사료된다. 또한 향후 제아잔틴이 어떠한 세포내 기전으로 LPS에 의해 염증이 유발된 피부를 개선하는지에 대해 추가연구 및 임상연구가 필요할 것으로 사료된다.
Acknowledgements
이 논문은 2019학년도 세명대학교 교내학술연구비 지원에 의해 수행된 연구임.
Figure 1.
Cytotoxicity of zeaxanthin in RAW264.7 cells.
RAW264.7 cells (4×104 cells/well) were seeded in 96-well tissue culture plates and treated with zeaxanthin for 24 h. The CCK-8 kit was used to perform the cell viability assay. These results are presented as mean±S.D. of the percentage of control optical density in triplicate. S.D., standard deviation. *value relative to the control (*p<0.05).
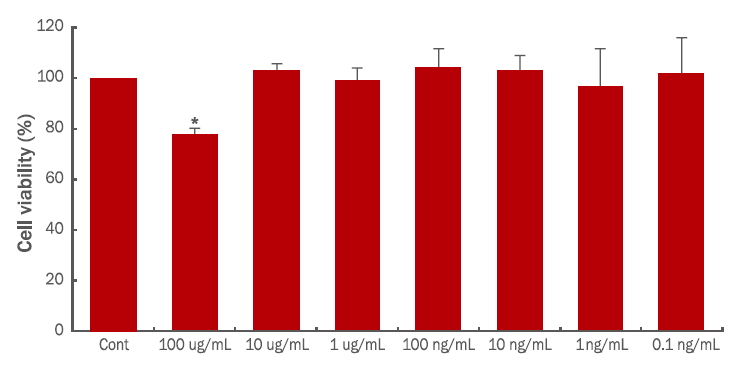
Figure 2.
Effects of zeaxanthin on NO synthesis in RAW264.7 cells.
RAW264.7 cells were seeded in 35 mm culture dishes and treated with zeaxanthin for 24 h. The media containing the cells were collected and analyzed for NO using a NO detection kit (A). Real-time PCR analysis of iNOS mRNA (B). The data represents the mean±S.D. of three independent experiments. NO, nitric oxide; PCR, polymerase chain reaction; LPS, lipopolysaccharide; S.D., standard deviation. *value relative to the control; **value relative to the LPS-treated condition; *, **p<0.05.
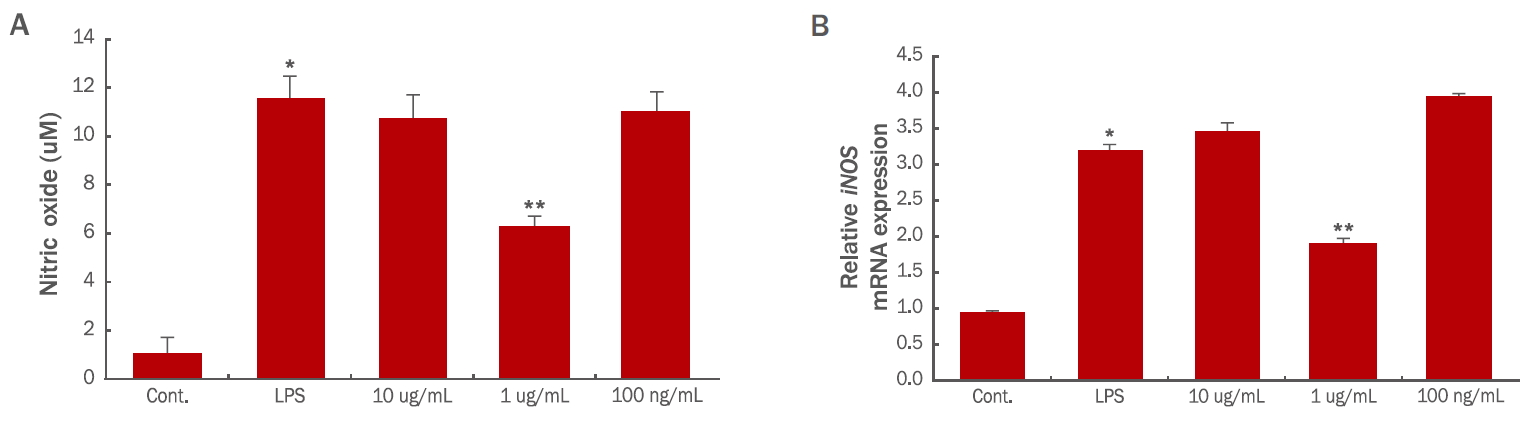
Figure 3.
Effects of zeaxanthin on PGE2 synthesis in RAW264.7 cells.
RAW264.7 cells were seeded in 35 mm tissue culture dishes and treated with zeaxanthin for 24 h. The media containing the cells were collected and analyzed for PGE2 levels by ELISA (A). Real-time PCR analysis of COX2 mRNA (B). The data represents the mean±S.D. of three independent experiments. PGE2, prostaglandin E2; COX2, cyclo-oxygenase 2; S.D., standard deviation. *value relative to the control; **value relative to the LPS-treated condition; *, **p<0.05.

Figure 4.
Effects of zeaxanthin on the expression of inflammatory cytokines.
Real-time PCR analysis of pro-inflammatory transcription factors was carried out for IL1α (A), IL1β (B), IL6 (C) and TNFα (D). The graphs are shown as mean±S.D. of three independent experiments. PCR, polymerase chain reaction; IL1α, interleukin 1 alpha; IL1β, interleukin 1 beta; IL6, interleukin-6; TNFα, tumor necrosis factor-α. *value relative to the control; **value relative to the LPS-treated condition; *,**p<0.05.
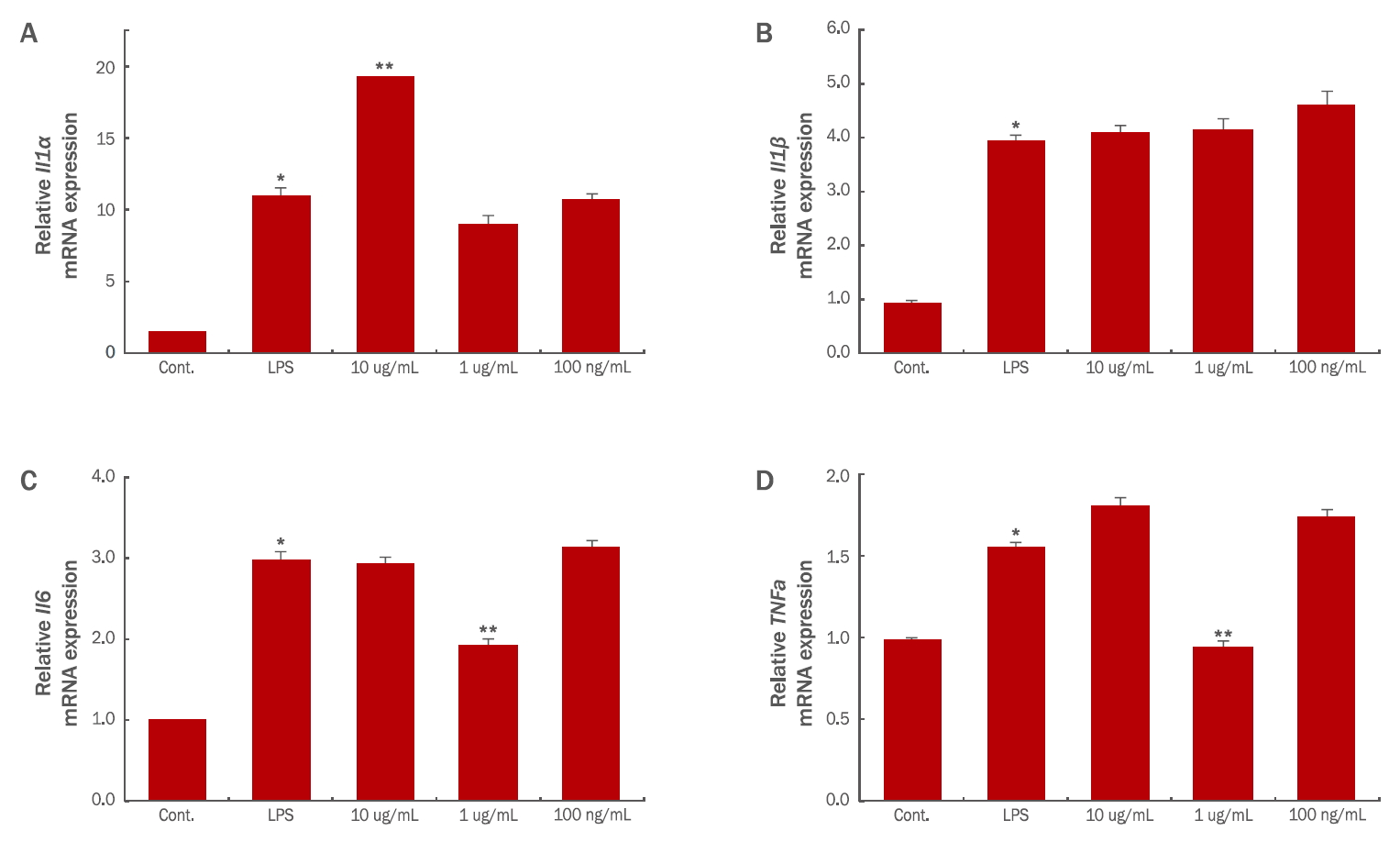
Table 1.
Gene name and assay ID number in real-time RT-PCR analysis
|
Symbol |
Gene name |
Assay ID |
|
NOS2
|
Nitric oxide synthase 2, inducible |
Mm00440502_m1 |
|
COX2
|
Cyclooxygenase 2 |
Mm03294838_g1 |
|
IL1α
|
Interleukin 1 alpha |
Mm00439620_m1 |
|
IL1β
|
Interleukin 1 beta |
Mm00434228_m1 |
|
IL6
|
Interleukin 6 |
Mm00446190_m1 |
|
TNFα
|
Tumor necrosis factor alpha |
Mm00443258_m1 |
|
GAPDH
|
Glyceraldehyde-3-phosphate dehydrogenase |
Mm99999915_g1 |
References
Akira S, Takeda K. Toll-like receptor signaling. Nature Reviews Immunology 4: 499-511. 2004.  Albina J, Reichner J. Nitric oxide in inflammation and immunity. New Horizons 3: 46-64. 1995.
Amar EC, Kiron V, Satoh S, Watanabe T. Enhancement of innate immunity in rainbow trout ( Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish and Shellfish Immunology 16: 527-537. 2004. Baek YM, Choi JY, Lee CW, Jeon YS, Han JT, Jang SI, Yoo HS. Effects of Chinemys reevesii on lipopolysaccaride-indused inflammatory reactions. Korean Journal of Oriental Physiology & Pathology 26: 26-34. 2012.
Ban JY, Kim BS, Kim SC, Kim DH, Chung JH. Microarray analysis of gene expression profiles in response to treatment with melatonin in lipopolysaccharide activated RAW 2647 cells. Korean Journal of Physiology Pharmacology 15: 23-29. 2011. Bartley GE, Scolnik PA. Plant carotenoids: pigments for photoprotection, visual attraction, and human health. The Plant Cell 7: 1027-1038. 1995.
Berry A, Janssens D, Hümbelin M, Jore JP, Hoste B, Cleenwerck I, Vancanneyt M, Bretzel W, Mayer AF, Lopez-Ulibarri R, Shanmugam B, Swings J, Pasamontes L. Pasamontes L. Paracoccus zeaxanthinifaciens sp. nov., a zeaxanthinproducing bacterium. nov International Journal of Systematic and Evolutionary Microbiology, 53: 231-238. 2003.
Bertram JS, Vine AL. Cancer prevention by retinoids and carotenoids: independent action on a common target. Biochimica et Biophysica Acta 1740: 170-178. 2005. Chew BP, Park JS. Carotenoid action on the immune response. Journal of Nutrition 134: 257S-261S. 2004.  Gabay C. Interleukin-6 and chronic inflammation. Arthritis Research & Therapy 8: 1-3. 2006.
Gomez PF, Pilllinger MH, Attur M, Marjanovic N, Dave M, Park J, Binagham CO, Al Mussawir H, Abramson SB. Resolution of inflammation: prostaglandin E2 dissociates nuclear trafficking of individual NF-kappa B subunits (p65, p50) in stimulated rheumatoid synovial fibroblasts. Journal of Immunology 175: 6924-6930. 2005. Hannibal L, Lorquin J, D’ortoli NA, Garcia N, Chaintreuil C, Masson-Boivin C, Dreyfus B, Giraud E. Isolation and characterization of canthaxanthin biosynthesis genes from the photosynthetic bacterium Bradyrhizobium sp. strain ORS278. Journal of Bacteriology 182: 3850-3853. 2000. Harker M, Hirschberg J, Oren A. Paracoccus marcusii sp. nov., an orange Gram-negative coccus. nov International Journal of Systemic Bacteriology, 48: 543-548. 1998. Harris SG, Padilla J, Koumas l, Ray D, Phipps RP. Prostaglandins as modulators of immunity. Trends in Immunology 23: 144-150. 2002. Hirohashi N, Morrison DC. Low-dose lipopolysaccharide (LPS) pretreatment of mouse macrophages modulates LPS-dependent interleukin-6 production in vitro. Infection and Immunity 64: 1011-1015. 1996. Horwood NJ, Page TH, McDaid JP, Palmer CD, Campbell J, Mahon T, Brennan FM, Webster D, Foxwell BM. Bruton's tyrosine kinase is required for TLR2 and TLR4-induced TNF, but not IL-6, production. Journal of Immunology 176: 3635-3641. 2006. Im DY. Volatile compounds analysis of the extract from dried bark of Prunus sargentii and physiological activity of the main compound, benzaldehyde. Asian Journal of Beauty and Cosmetology 12: 155-162. 2014.
Jones DL, Rando TA. Emerging models and paradigms for stem cell ageing. Nature Cell Biology 13: 506-512. 2011. Kim J, Lee CW, Kim EK, Lee SJ, Park NH, Kim HS, Kim HK, Char K, Jang YP, Kim JW. Inhibition effect of Gynura procumbens extract on UV-B-induced matrix-metalloproteinase expression in human dermal fibroblasts. Journal of Enthnopharmacology 137: 427-433. 2011. Kim S, Kang BY, Cho SY, Sung DS, Chang HK, Yeom MH, Kim DH, Sim YC, Lee YS. Compound K induces expression of hyaluronan synthase 2 gene in transformed human keratinocytes and increases hyaluronan in hairless mouse skin. Biochemical and Biophysical Research Communications 316: 348-355. 2004. Kirkwood TB. Understanding the odd science of aging. Cell 120: 437-447. 2005. Kurihara H, Koda H, Asami S, Kiso Y, Tanaka T. Contribution of the antioxidative property of astaxanthin to its protective effect on the promotion of cancer metastasis in mice treated with restraint stress. Life Science 70: 2509-2520. 2002. Lawrence T, Willoughby DA, Gilroy DW. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nature Reviews Immunology 2: 787-795. 2002. Lee HJ, Sim BY, Bak JW, Kim DH. Effect of Gamisopungsan on inflammatory and DNCB-induced dermatitis in NC/Nga in mice. Korean Journal of Oriental Physiology & Pathology 28: 146-153. 2014.
Lord PC, Wilmoth LM, Mizel SB, McCall CE. Expression of interleukin-1 alpha and beta genes by human blood polymorphonuclear leukocytes. The Journal of Clinical Investigations 87: 1312-1321. 1991. Mayne ST. Beta-carotene, carotenoids, and disease prevention in humans. FASEB Journal 10: 690-701. 1996. Namazi MR. Cetirizine and allopurinol as novel weapons against cellular autoimmune disorders. International Immunopharmacology 4: 349-353. 2004. Noman AS, Koide N, Hassan F, I-E-Khuda I, Dagvadorj J, Tumurkhuu G, Islam S, Naiki Y, Yoshida T, Yokochi T. Thalidomide inhibits lipopolysaccharide-induced tumor necrosis factor-a production via down-regulation of MyD88 expression. Innate Immunity 15: 33-41. 2009. Roitt IM, Brostoff J, Male DK. Immunology sixth edition. Mosby. London. pp119-441. 2001.
Shon MS, Song JH, Kim JS, Jang HD, Kim GN. Anti-oxidant activity of oil extracted from Korean red Ginseng and its moisturizing function. Asian Journal of Beauty and Cosmetology 11: 489-494. 2013.
Talwar HS, Griffiths CE, Fisher GJ, Hamilton TA, Voorhees JJ. Reduced type I and type III procollagens in photodamaged adult human skin. Journal of Investigative Dermatology 105: 285-290. 1995. van Triel JJ, Arts JH, Muijser H, Kuper CF. Allergic inflammation in the upper respiratory tract of the rat upon repeated inhalation exposure to the contact allergen dinitrochlorobenzene (DNCB). Toxicology 269: 73-80. 2010. Yun KJ, Kim JY, Kim JB, Lee KW, Jeong SY, Park HJ, Jung HJ, Cho YW, Yun K, Lee KT. Inhibition of LPS-induced NO and PGE2 production by asiatic acid via NF-kB inactivation in RAW 22647 macrophages: possible involvement of the IKK and MAPK pathways. International Immunopharmacology 8: 431-441. 2008. Zhang Y, Ramos BF, Jakschik BA. Neutrophil recruitment by tumor necrosis factor from mast cells in immune complex peritonitis. Science 258: 1957-1959. 1992.
|
|












